4 / 2019 / vol. 8
Kosmetologia Estetyczna
453
ARTYKUŁ NAUKOWY
KOSMETOLOGIA ESTETYCZNA
N
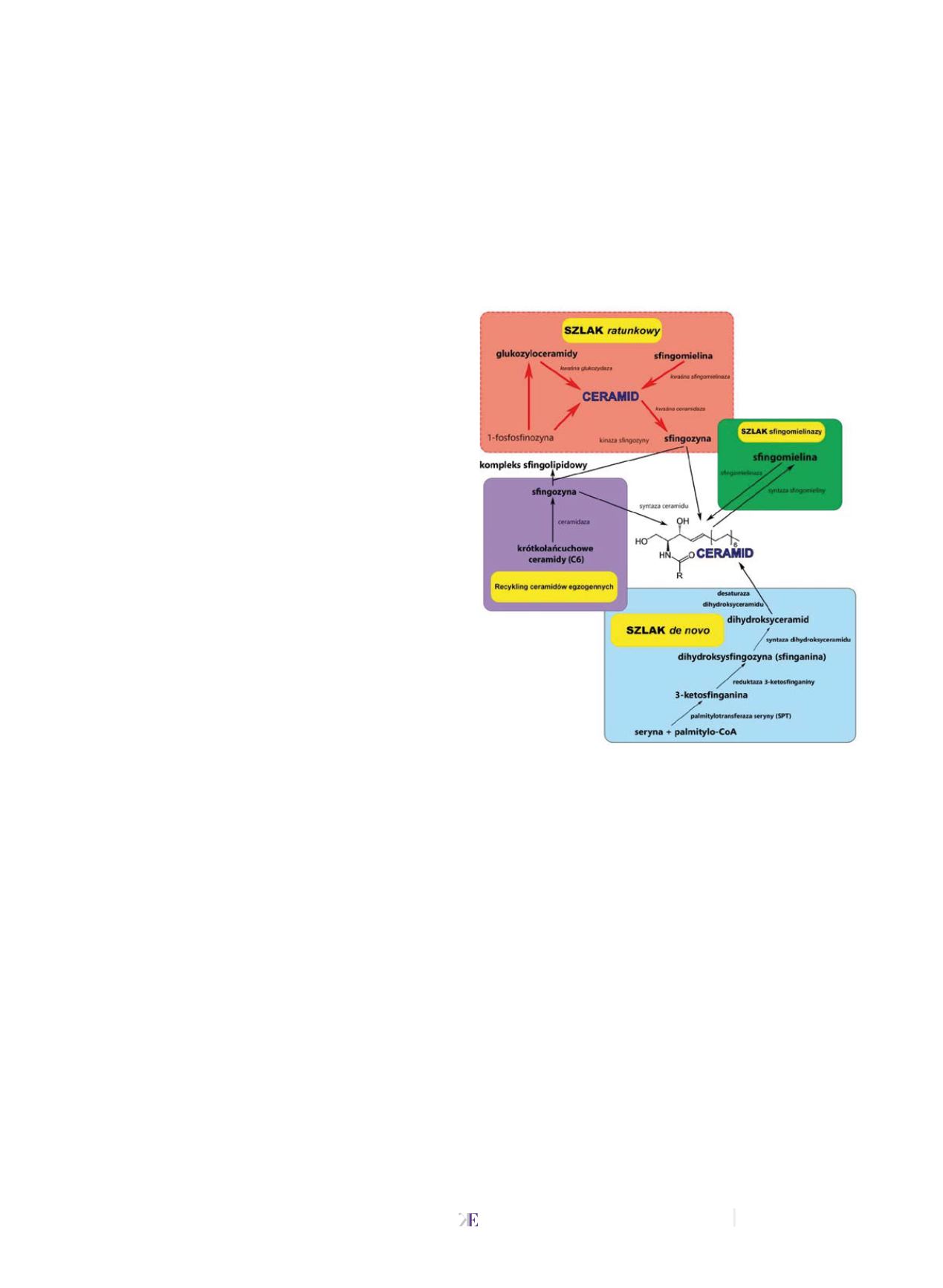
Znanych jest 6 różnych syntaz ceramidu, różniących się po-
winowactwem do określanych kwasów tłuszczowych oraz
dystrybucją tkankową. Syntaza 3 jest odpowiedzialna za nie-
zwykłe ceramidy skóry bowiem wykorzystuje aktywne kwa-
sy tłuszczowe o dwudziestu sześciu atomach węgla (C26-CoA)
i także dłuższe. Natomiast syntaza 4, również występująca
w skórze użytkuje krótsze łańcuchy węgla występujące w ak-
tywnych kwasach (C18 do C22-CoA) [12-20].
Degradacja ceramidów
do sfingozyny i kwasów tłuszczo-
wych odbywa się za pośrednictwem ceramidaz. Do tej pory
zostało zidentyfikowanych pięć różnych enzymów katalizu-
jących tę reakcję: kwaśna ceramidaza, obojętna ceramidaza
i trzy izoformy zasadowej ceramidazy. Podobnie jak syntazy
ceramidu, ceramidazy różnią się powinowactwem do kwasów
tłuszczowych i miejscem występowania w komórce. Dla kwa-
śnej ceramidazy specyficznymi substratami są krótko (C≤6)
i długołańcuchowe (C≥16) ceramidy. Obojętna ceramidaza
natomiast preferencyjnie katalizuje reakcje z udziałem długo-
łańcuchowych ceramidów. Wykazano
in vitro,
że gwałtowny
wzrost aktywności zasadowych ceramidaz powodują jony
wapnia. Te ceramidazy katalizują reakcje z udziałem cerami-
dów o długich (C≥24) łańcuchach węglowych [18-27].
Nie stwierdza się w komórkach procesu akumulacji określo-
nych ceramidów. Aby zachować ich homeostazę, dochodzi do
transportu ceramidów z siateczki śródplazmatycznej do apara-
tu Golgiego, gdzie stają się prekursorem innych sfingolipidów,
takich jak sfingomielina i glikosfingolipidy. W tym transporcie
ważną rolę odgrywa specjalne białko cytozolowe wykazujące
specyficzność substratową co do długości łańcucha węglowe-
go, jak również stereospecyficzności
.
Białko to, najefektywniej
transportują ceramidy o następujących długościach łańcucha
węglowego: C14, C16, C18 i C20.
•
Ceramidy powstają także w ramach tzw.
„cyklu sfingomie-
linowego”
(zielony, rys. 2). Wówczas w wyniku hydrolizy
sfingomieliny w obecności odpowiedniej sfingomielinazy
powstaje ceramid. Niekorzystnie działające bodźce komór-
kowe np. szlak indukowany stresem, czy infekcja, powodują
hydrolizę sfingomieliny w wyniku aktywacji sfingomielinaz.
Klasyfikacja tych enzymów opiera się na różnicach optimum
pH katalizowanej reakcji i w związku z tym wyróżnia się:
sfingomielinazę kwaśną, obojętną oraz alkaliczną. Sfingomie-
linaza kwaśna znajduje się wewnątrz komórki i wykazuje
najwyższą aktywność w pH 4,0-4,5 natomiast obojętna znaj-
duje się po zewnętrznej stronie błony plazmatycznej i wyka-
zuje optymalną aktywność w pH 7,4 oraz wymaga obecności
jonów magnezu (Mg
+2
) lub manganu (Mn
+2
)
jako aktywatora.
Badania wykazały, że degradacja sfingomieliny w wyniku
aktywacji kwaśnej i obojętnej sfingomielinazy trwa kilka se-
kund/minut w porównaniu do syntezy
de novo
ceramidów,
której czas trwania wynosi kilka godzin [17-24].
•
Trzeci szlak
(pomarańczowy, rys.2) pozyskiwania cerami-
dów w tzw. szlaku ratunkowym zachodzi w późnych endo-
somach i w lizosomach, gdzie katabolizmowi ulegają złożone
glikosfingolipidy mające istotne znaczenie w budowie błon
biologicznych.
Uwolnione ceramidy przy udziale kwaśnej
ceramidazy przekształcane są do sfingozyny. Z badań wy-
nika, że sfingozyna opuszcza lizosomy i może być metaboli-
zowana do ceramidów lub ulegać fosforylacji do sfingozyno-
-1-fosforanu [25-28].
Rys. 2
Szlaki powstawania ceramidów
Źródło:
[32]
POCHODZENIE LIPIDÓWWARSTWY ROGOWEJ
Lipidy warstwy rogowej są syntetyzowane głównie w naskór-
kowych komórkach posiadających jądra i są gromadzone w ich
organellach o ograniczonej liczbie błon zwanymi ciałkami blasz-
kowatymi lub lamelarnymi. Ciałka lamelarne zawierają prekur-
sorowe lipidy w skład których wchodzą głównie glukozyloce-
ramidy, sfingomieliny, cholesterol, fosfolipidy oraz odpowiednie
enzymy hydrolityczne. Zawartość ciałek ulega egzocytozie na
granicy warstw ziarnista-rogowa do przestrzeni międzykomór-
kowej i tam poddawana jest działaniu enzymów. W tym pro-
cesie wymagane jest co najmniej jedno białko transportujące
należące do rodziny ABC typu ABCA12, które jest ulokowane
w granicznej błonie ciałka lamelarngo. Fuzja ciałek z błoną pla-
zmatyczną w najwyżej zlokalizowanych jądrzastych komór-
kach naskórka (
stratum granulosum
), umożliwia przetwarzanie
prekursorów lipidów w domenie pozakomórkowej [25].


